Living creatures have, during their evolution, acquired an implicit knowledge of their environment. They have, by selective pressures in the flow of many generations, been shaped to respond sensibly to their usual environment. This biological process has led to phylogenetic (ancestral) cognition. Cognition is expressed in the body-forms and -functions that exactly fit their environment:
By using the word 'cognition' to stand for 'knowledge embodied in structure and functions', we borrow from the sciences of the mind. Cognition is ordinarily used for mental processes such as perception, memory and thinking in the higher primates. In the present context cognition is used in the broad biological sense of "wisdom of the body", as expressed in its form and function, and as acquired in the course of evolution by the Genetic System for Adaptation and Defence (GAD). The meaning of cognition is thus transferred from the time-scale of human life events (minutes - years) to that of the evolutionary time-scale (centuries-aeons). J. Lovelock (1988) has gone even further and has described the entire biosphere, including the planet Earth itself, as one living organism (Gaia) with a cognitive network that touches and includes everything.
Extending the meaning of the term 'cognition' is a step towards reintegrating the sciences, and should be recommendable for that reason. Another example of extending the meaning of a word is the use of 'learning' for 'improved responding to environmental challenge on the basis of experience'. We are used to think that learning takes place in the brain. The immune or lymphoïd system for adaptation and defence (LAD) however is as such a complete learning system on another level. It has a memory that is extended by experience, and it keeps its internal representation of the environment up to date by adding to its knowledge. It also has a value system: an immune system that enables the individual to assess its material environment and so to interact with it for food and protection. In cooperation with the nervous system it discriminates what can be taken in and what is to be avoided.
In the course of evolution plant and animal species are forever improving their means to protect their integrity, to maintain and multiply themselves. The learning process in the evolutionary time scale is augmented by learning in the lifetime of the individual animal (ontogenetic learning). The difference between plants and animals in this respect is that plants have too little memory storage to learn much during one single lifetime; however they learn relatively fast in the evolutionary time scale, as appears from the abundant varieties of wild and cultivated plants.
Learning in an individual animal's life time resembles evolutionary learning, except for the time-scale. In immune and neural learning, Darwinian processes of variation and selection take place at a rapid rate. The generations of lymph-cells and neuronal components follow each other at a high rate, so that selections can take place at short intervals. Immune globulins are produced by the millions every second. The rate of learning is immense: the immune system (LAD) responds to change a million times and the nervous system (NAD) even a billion times faster, than the response rate in the time-scale of evolution of an animal species.
The two systems within the vertebrate body that are open to evolutionary processes i.e. the lymphoid and the neural system, develop their adaptive memory-content during the individual's lifetime. In this way they foster appropriate behaviour in response to new stimuli. Such is the surprising solution of the enigma of learning: by varying the cellular and humoral output and selectively reinforcing the most fitting elements the best adaptive responses are fostered. The lymphoid system and the neuronal system belonging to one and the same organism, form part of each other's environment and as such influence each other's path of learning.
That these two systems work closely cooperate is the more fascinating since they are so different in form and substance. The immune system is for the greater part fluid, although thymus and bone-marrow and spleen provide a kind of substance; the brain is for the greater part solid. During the foetal stage nerve cells can migrate freely from one place to another, directed by similar fields of attraction as exerted by the lymphoid cells of the immune system. During it's whole life the brain restructures it's information content, even more freely than the lymphoid system does.
The two systems, LAD and NAD, enable individuals to regulate their interactions with the environment. Although the central nervous (neural) system responds a thousand times faster than the immune (lymphoid) system both systems have many properties in common:
Some messengers in the immune system have a relatively short response time, that comes near the time-window of slow-acting neural components belonging to the autonomous system. Because systems that oscillate in closely corresponding frequencies influence each other, the immune and nervous systems interact, via hormones and other pathways of the autonomous system. A similarly mediated interaction is discussed with relation to epigenesis and metabolism in chapter (3) on Embryogenesis.
Throughout the vegetal and animal kingdoms chemical signals are active, as internal messengers or as messengers between plants and animals, as attractants or repellents. In higher animals and man the "chemical" senses remain active in receiving and transmitting vital information. Tastes and smells are detected by scanning and recognising the molecular surface configurations of odorants, in a similar way as antibodies recognize antigens. Recognition leads to depolarisation of neurons in the olfactory epithelium. The signals are decoded by the "rhinencephalon", a phylogenetically ancient drive and motivation centre that is a part of the limbic system of the brain's cortex. Odours can act as pheromones, also in humans. They often give rise to strong motives and to immediate, partly involuntary action.
Before we start studying the maps of the above mentioned systems that have specialised in learning (the lymphoid and the neural system) I want to account for the fact that these maps have a spherical form, and are divided in concentric zones that envelop each other.
The logic of evolution comes to the surface: create ever more complex assemblies in synergy with the environment (4.3). This is called differentiation (Map 5.2.1). The first clusters of cells in an early evolutionary stage were still rather homogeneous, like the amoebe's cell mass (3.1). In the course of evolution tissues differentiated, organs specialised in digestion, motion, defence etc.
Concentricity is a device that reduces the complexity of the organism without resorting to an overly simple one-way hierarchy (Yakovlev 1970). In a concentric organisation the hierarchy goes both ways: from the inside outwards, and from the outside inwards. The central structures respond to environmental change with a long time-constant and impose their standard values on the peripheral layers. These have shorter time-constants, which enable them to respond rapidly to the environment while implementing the instructions coming from the central core.
Central structures have the following properties:
Surface structures on the contrary show
When a core structure detects an incongruity with respect to its standards, it dispatches a message to it's neighbouring peripheral structure, which in turn starts operations to diminish the incongruity. The operant response needs some time before it will have reduced it's motivating signal; the higher the oscillating frequency of the corrective or regulatory circuit is, the shorter the time interval will be. Other oscillating circuits that also interact with the motivating parameter (the messenger of incongruity) will compete for effectiveness with the first circuit. The one that has the best adaptive effect will be the one most likely to be selected and to subsequently become a permanent part of the system.
Another result of learning is anticipating a need for adaptive action. Any change that is recognisable as a stimulus can, when it concurs with a message, become associated with that message. From then on, the message predicts the stimulus and sets the corrective actions in motion before the incongruity is actually noticeable. This is classical conditioning, or respondent conditioning. By this learning process the NAD learns avoidance and appetitive behaviour. It enables the individual to avoid impending negative (hostile) stimulation and to seek positive (feeding) stimulation from the environment.
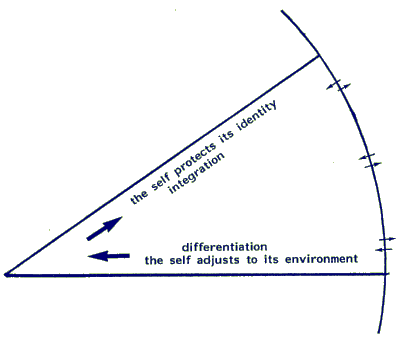
To protect themselves living systems, from the most elementary to the most complex forms, employ two strategies that complement each other:
(a) adjustment: when a living system is challenged by its environment it responds by adjusting to the challenge. A succession of challenges exerts pressure for continuing adaptation and directs differentiation and increased complexity of function and form
(b) maintaining identity: the genetic system has set limits to the possibilities for change; individuals protect their 'self' and maintain their specific and individual identity against pressure to adjust to the environment.
The opposing tendencies for change (a) and resistance to change (b), are in continual dialogue. The synergy, finally reached by the pull of opposing fields, forces optimal growth and learning. The tendency to change is necessary to optimally adjust to environmental requirements, such as protection against climatic change or a change in predator population. The genetic system offers a vast range for adjustment and individual learning. It also offers resistance to change because there is a limit to the range of genetic potential. This limit guarantees that eventual adjustments are in agreement with the master plan of the individual. The genome guards against weakening of identity by over-adjustment, which would lead an individual to disintegrate (as after repeated and severe emotional traumatisation, when an individual is compelled by force to forsake his identity).
Fig. 5.2.1 (at begin of this paragraph) represents the process of synergy. It summarizes the dialogue of nature (heredity) and nurture (environment) in development and learning. Because of its explanatory power, a sphere with concentrical layers is the appropriate model to represent the continuous dialogue of centripetal (nurture) and centrifugal (genetic) information pathways. The dialogue and the resulting synergy is characteristic for immune and neuronal learning within individuals. A spherical model with a core and various layers or growth-zones will answer such questions as: how an individual meets environmental challenges, how he learns to anticipate positive and negative events. For the sake of simplicity our three-dimensional sphere has been reduced to a flat disk. It is not even a full disk, only a partial cross-section of the sphere of a living system is shown, in the form of a sector, which leaves the rest of the disk invisible.
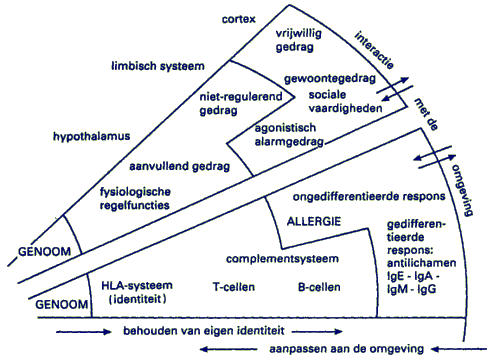
Map 5.2.2: Two learning systems
We see a curved surface: the boundary between the organism and its environment. It is the "surface-membrane" where the exchange of information between the individual and its environment takes place. At this level the LAD deploys its antigen sniffing lymphocytes, and the NAD its sensory scanning organs: smell, sight, touch, hearing. Toward the left of the diagram, the sides of the sector converge in the hub of the sphere. There, far removed from the surface, is the location of the inherited factors, which mark the genetic identity of the individual. It is called the genome and its material basis is the chemical building-code laid down by the DNA molecules. The genome constitutes the evolutionary memory and maintains the individual's identity against pressure coming from the environment. The continuous information issuing from the genome protects the individual against over-adaptation and the disintegration that would be the result.
The genome is the hard core of the individual that resists change and only adapts under long-term pressures in the evolutionary time scale. Fortunately it leaves room for short-term adaptations which we label as epigenetic or ontogenetic learning. Information about the environment, detected at the surface, is carried inwards where it meets the outwards directed information issuing from the genome. Genetic information is carried outwards to the external layers, where it engages in a "dialogue" with messages received from the environment. The reconciliation resulting from the interaction of opposing information streams is called learning. This may be the acquisition of new and meaningful cognition, or more differentiated skills, both of which will contribute to profitable adjustment while safe-keeping the individual's identity. On a global level, adaptive changes seem to be "instructed" by the environment; on the molecular level they turn out to be "selected" (Jerne 1967). In order to understand this, we'll turn once more to the immune system, since its development is exemplary for a learning process.
Evolution of an animal species (phylogenesis) is a learning process on the time-scale of aeons. What is usually called 'learning' occurs during the lifetime of one individual (ontogenesis). Learning by the immune system consists in selecting the most appropriate antibody response to an environmental challenge. Acquiring (learning) the best fitting immune response to a new antigen, occurs in a time-scale of days/weeks.
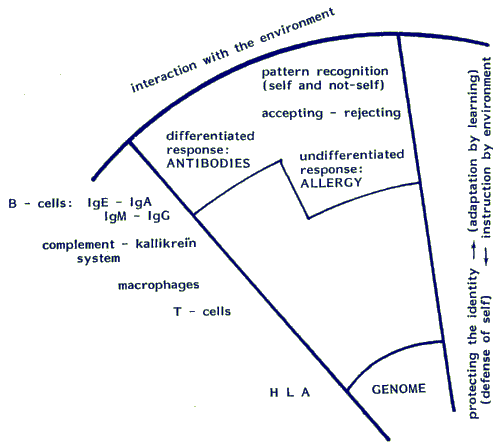
Map 5.2.3: The lymphoid system for adaptation and defence
Map 5.2.3 is phylogenetically arranged: components of the immune system that have been formed early in the evolutionary history are close to the centre of the sphere. The core systems are surrounded by ancillary subsystems and these are resistant to change. Humans share the same subsystems with many other vertebrate species. Closer to the surface, in interaction with the environment, are highly differentiated components of the system: the antibodies. They are phylogenetically younger and more adjustable. Several types of immune-globulins (antibodies) that are active in man, are not found in the lower vertebrates. The map also shows some aspects of ontogeny: at birth only the central part of the sphere is fully developed. The more peripheral zones mature and differentiate in the course of a learning process, that takes place in blood-forming organs, thymus and lymph-nodes during early childhood. (The relation of growth, maturation and learning is explained in Map 9.5.1)
The lymphoid or immune system is a refined system for chemical recognition of the surrounding world. Its function can be defined as supplying the young individual with an up to date internal image of the environment in which it is growing up. Since every individual has it's own specific history of encounters, each resulting image differs from anyone else's. Relevant stimuli from the environment are called antigens or immunogens, since they give rise to an immune response, i.e. secretion of antibodies. Every meeting with an antigen is recorded and stored as a memory trace in the network of lymphocytes and the proteins they produce. Antigenic properties are also labelled in a value system that discriminates between self and non-self, and this label is attached to the record. When confronted later, and in another context with a familiar antigen, the system will decide if the stimulus is in the self or non-self category and whether it is acceptable (to be approached) or unacceptable and to be avoided. It does so by continuously checking antigens against the individual's identity-tags: checks are carried out to see if the component can be assimilated or should be rejected.
Most of the relevant information is coded in the form of surface configurations on antibodies, i.e. macromolecular proteins belonging to the class of immune globulins (Ig's). They circulate freely, or remain on the surface of the lymphocytes that produce them. Lymphocytes (white blood cells) of various types are produced in bone marrow. Their rate of production is facilitated or inhibited, depending on the antigenic properties of the surface structures which
they carry with them. When a lymphocyte finds a good match with a relevant antigen it's production is selectively reinforced: the bond between stimulus and receptor is the signal which starts reproduction. Antibodies of various types also recognize each other, as if they were themselves antigens. Thus they facilitate or inhibit the growth rate of their respective parent lymphocytes. As a result, a network is set up and maintained, in which oscillating population densities keep each other in check. The wave-propagation of population-increase and -decrease is similar to the one described in 3.1 to 3.3.
|
In this way the spatial properties of antigens and antibodies - surface configurations - are transfigured into temporal properties within the system - coupled oscillations in a network -. Pringle (1951) has discovered that coupling of non-linearly oscillating population densities, and selection of the oscillations that best serve life and health, is the basis of learning. His discovery has been worked out for the neural system (NAD) by Thorpe (1963) and Grossberg (1982). The kinetics of the lymphoid network (LAD) present a promising area of research. The symphony of oscillations carry the essential meaning, not the cells that produce them, just as music is a message, independent of the instruments that produce the oscillations in the air. |
N.K. Jerne (1985) has compared the endless combinatory possibilities of meaningful configurations on the surface of antibody molecules with words and sentences of a language. Just as the LAD generates an appropriate response to molecules that the system has never met before, so the VAD (language) can formulate, in a novel combination of syllables and phrases, meaningful expressions that have never before been used. This analogy between language and the immune system drapes the LAD with a poët's cloak.
Summing up the function of the lymphoid system, one can say that the lymphocyte network keeps its image of the material world up to date. It educates itself continually by scanning the internal and external environment for familiar and unfamiliar features. The information is processed (evaluated and labelled) and passed on to the deeper structures for permanent storage. A schematic representation of components of the lymphoid system is given in Map 5.2.3 They are arranged in spherical form around the genome. The standard or yard-stick for compatibility of antigens is a complex of features in the genome: human lymphocytic antigen complex (HLA) or, in more general terms, the major histocompatibility complex or MHC. This is "given" and does not change in the individual's life time. It can only be changed in future generations by selective mating of the individuals who carry the genome.
Less rigid and more adaptable are other parts of the lymphoid system situated closer to the periphery of the sphere. They form networks as described, and are in immediate contact with the environment, performing such executive tasks as scanning, recognizing and tagging antigens for rejection or assimilation.
People differ widely as to their sensitivity for antigenic stimulation and their immune responsivity. One condition for acquiring an adequate immune response is that the range of antigenic stimulation be within the limits of an individuals' coping capacity. If the dose of antigen is too low, discriminatory power, recognition and adequate response will fail to develop. The stimulation is recorded, to be sure, but not as a challenge. In consequence false tolerance may develop (non-response). This may cause problems later: when the stimulus ever returns in massive quantity, there is no adequate answer and the individual is left defenceless.
Likewise if the dose is too high the stimulation will not lead to the development of discriminatory power either. When an overwhelming challenge presents itself, the defence system cannot but fall back on violent primitive reactions that defeat any attempt at a differentiated recognition-response (regression). A meaningful feature of the Map 5.2.3 in this respect is the enclave labelled "allergy". It represents a set of non-specific defence-reactions, more primitive than the differentiated antibody responses in the outer layers of the sphere. As a tool for emergencies this intermediate layer is activated when the lymphoid system has not yet learned how to cope with the allergenic substance in a more appropriate way, or when the system is being flooded with larger quantities of the antigen than it can handle.
Through the immune or lymphoid system the individual communicates with its environment. The system, with cells spread through all tissues of the body, is a sense-organ with a memory attached that discriminates between self (the body's own components) and non-self (foreign matter and organisms in the environment). It defends the self and eliminates non-self substances. Throughout infancy the system learns to know and to recognize the usual antigens that occur in the environment where it grows up. The immune system works with macromolecules that are called antibodies. Antibodies are produced by lymphocytes. The lymphocytes and the freely circulating antibodies have recognition sites on their surfaces by which they can identify, and discriminate between thousands of different organic and non-organic substances that occur in nature. In the early years of childhood the immune system succeeds in cultivating a network of recognition sites that accurately represents the environment in which the individual grows up, just as the brain develops an internal image of the smellable, touchable, audible, visible world. This is made possible by Darwinian selection in a fast time-window (with a response time of days/weeks) that is even faster than the evolution of microorganisms. The shapes of antibody molecules have been selected by the antigens which the individual encounters. When a lymphocyte produces an antibody with recognition sites on it's surface that fit the antigen fairly well it's production is stimulated more than that of a lymphocyte generating a less well fitting antibody. So the best recognition sites increase in numbers and the other varieties perish and disappear from the scene. It is a Darwinian type of selection, not unlike the breeding of dogs with special properties of shape and behaviour. Selective breeding of antibodies however takes place within the individual's life time.
The immune system's molecules and cells not only recognise antigens, they also recognise each other. Since all recognised objects are called antigens, the immune-globulins have a double role, that of antigen and of antibody. As a consequence, antibody producing lymphocytes can stimulate as well as suppress other cells of their kind. By engaging in mutually suppressing and stimulating activities, the numerous varieties of individual cells and molecules constitute a network, a response-system of interacting components, an "evolving species". These oscillations are comparable to those in a "prey - predator"relationship.
When the immune network is stimulated by an antigen this elicits an increase of those lymphocytes that secrete the best fitting type of antibody for that specific antigen. There is some time-delay due to the succeeding generations of lymphocytes in the evolution of the immune response. In due time, the encounter with an antigen produces a shock-wave that travels through the system and that eventually leads to the elimination of the antigenic stimulus (by phagocytosis, agglutination and lysis or other means). After a while the shock wave (and the accompanying fever and malaise) dies down and the equilibrium of the system is reestablished. One thing has changed however: there is a permanent memory trace of the disturbance in the form of circulating lymphocytes that preserve the internal image of the encounter with the antigen. At the next contact with the antigen the response will be fast and efficient, so that only a ripple of a disturbance will pass through the network, no fever, no malaise: the individual has become immune to that particular antigen. The lymphoid system has acquired an internal image of a relevant part of the perceived world, and this we have labelled cognition.
During their early years infants have learned to cope with foreign stimulation, such as different kinds of food and various infectious agents. "Foreign" has become more or less "familiar" in the process. A flexible response system has acquired a file of substances carrying the label of self (familiar) or non-self (foreign). After a successful learning period of several years the immune system has become able to function adequately.
Like any regulatory system the lymphoid network that maintains an equilibrium between self and non-self is never completely at rest. Even during periods of relative calm the constituents of the immune system will oscillate around mean values of population density. The coupled oscillators form an internally coherent dynamic network that responds more efficiently to stimulation than if it were in static equilibrium.
With advancing age the LAD becomes prone to errors. Inaccuracies in recognising and labelling lead to attacks on innocent organs in the category of self. This is the cause of auto- immune diseases which may affect the blood-forming organs, the lining of the joints, and the white matter of the nervous system. The phenomenon of auto-immunity has its counterpart in the VAD, where semantic confusion may lead to false alarms and unjustified verbal aggression.
Like the lymphoid system the nervous, or neuronal, system has evolved to carry out the following functions:
It is formed on the basis of emotional and sensorimotor experiences. To this end it has developed two different memory-tracks i.e. [1] the capacity for rapid attentional integration (short time memory), and [2] a buffer storage capacity (long term memory, Blumenthal 1977).
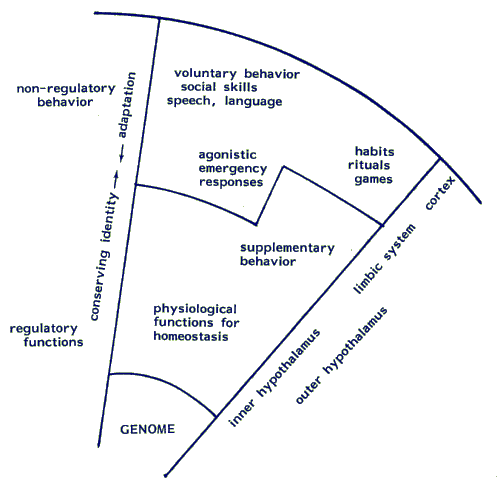
Map 5.6.1: The neural system for adaptation and defence
As in the map of the lymphoid system (5.2.3) we recognise in the NAD (Map 5.6.1) a concentrical arrangement of functions and behaviours: vital regulatory functions are close to the centre of the sphere, the more differentiated functions, such as voluntary behaviour and social skills, closer to the surface. The functions near the centre are rather rigid and hardly influenced by learning, closer to the periphery there is increasing plasticity. Most functions and behaviours are learned at an early age. As a consequence, the "behavioural style" with which an individual proceeds through life, has been formed in the first years of it's existence.
Most forms of behaviour are acquired by learning. But heredity determines the potential of an individual and sets limits to what it will be able to learn ('nature'). The kind of education ('nurture') as well as decisive incidents in the life history determine how much of the potential allowed by nature will be actualised. During childhood the environment selectively reinforces those elements from the developmental potential that fit the requirements of the moment. This is often a short-sighted strategy which may turn out to be maladaptive in the long run. Normally maladaptive behaviour will be replaced, before adulthood is reached, on the strength of more and better information and larger aims.
As long as it is alive the brain is engaged in 'adaptive learning': new connections between neurons are made, old ones are replaced. The neurons and their adjacent glia (supporting cells) constitute functional units. Their main business is combining or isolating streams of information. Combining takes place at the surface of cell-bodies and dendrites, the isolated transport takes place in the axones, that are part of the neuron cell and can extend from the spine to a toe. By changing its network properties, in interaction with the environment, the neural system is continuously improving its efficiency. Some parts of the brain are more flexible than others in this respect. Most flexible are those parts that have been acquired late in the phylogeny of man: the neocortex of both hemispheres or neo-encephalon. It surrounds the older parts of the brain: the paleo-cephalon and the spino-medullary part of the central nervous system.
- The 'hard' core of the brain is the substrate for innate reflexes that are resistant to modification by experience. It is immediately related to the internal maintenance of water-balance, blood-sugar levels etc.
- The midbrain together with the limbic system ('paleocortex') provide the substrate for the regularly recurring activities, concerned with alimentation and reproduction. When the demands on the physiologic systems exceed their capacity for control, supplementary behaviour maintains body-temperature, sustains water- and food intake. Closely related are appetitive and avoidance behaviours, sexual, fight, flight and withdrawal activities. Such emergency behaviours are called 'agonistic', they secure the integrity of the individual in extreme circumstances. In so doing they may interfere at unexpected moments with already acquired more sophisticated behaviour. This fact will prove to be of considerable interest for explaining otherwise inexplicable human communication disorders (Chapter 10). Although they are instinctive and inborn, their mode of execution is nevertheless largely modifiable by experience. The limbic system is considered to be the matrix for emotional and motivational learning. Early 'agonistic' survival strategies (neuroses) are stored here.
Farther away from regulatory processes we find a layer of skilled escape and avoidance behaviours, exploration and manipulation. They serve to improve the circumstances under which the individual tries to keep his physiological and emotional balance. Far removed from the physiologic homeostatic drives are the varied and voluntary behaviours. They are only loosely connected to the physiologic core functions and show a less stereotyped pattern than those more closely related to the physiological needs.
Regulatory behaviours are controlled by strictly defined substrates in and around the hypothalamus. The location of motivation for more voluntary behaviour, however, is not so strictly defined. We have an idea of how selecting an appropriate response works in a given instance. When a complex stimulus is received, it is rapidly scanned for cues that stand out because of their 'attention-value'. These are either general markers that are valid for all members of the species, or individual markers for which a person has been sensitized in particular. Incoming messages are then analysed in more detail on a higher level of the central nervous system, in the neo-encephalic part of the brain's cortex, where variable adaptive behaviours are organised. Variation in behaviour is wide, although always limited by the genetic potential of the individual. Within this limit, a selection from a large repertoire is made in response to immediate needs. Plasticity of the neo-encephalon allows the development of speech and language skills, in interaction with the environment. This is so in most humans, in some instances a genetic defect (e.g. a chromosomal aberration) or brain damage at birth limits the degree in which development of adaptation and adequate defence are possible.
From the foregoing we conclude that here is a sequence through which the genetic information passes in order to be expressed in one of the functional layers:
Physiological functions (deep structures) and voluntary behaviour (surface structures) supplement each other. An every-day example will illustrate this. When you are working in the sun, your autonomic system adapts to the situation by increasing the blood circulation to the body's surface and by sweating. When the need for heat-loss surpasses the capacity of your physiological regulation system, voluntary action will supplement it. By taking off coat and shirt or by taking a rest in the shade you carry out an imperative dictated by your physiology. It is easy to think of other examples of seemingly voluntary behaviour, that stem primarily from physiological drives: having breakfast, drinking a glass of water, going to sleep etc.
Cognitive processing not only serves to immediately carry out behaviour inspired by drive-motivated needs, it also anticipates physiological needs before they become too demanding. It is also inherent in humankind to freely play around with cognitive agility. As human beings we can conjure up possible and impossible worlds just by thinking. Language is a very suitable instrument for this creative activity, by which we try plan actions for ourselves and communicate about them with other people.
With our immune system we create an internal image of the material world in which we live so we can anticipate its challenges. As the syntax and grammar of the lymphoid network reveal themselves to the experts in the field, they begin to see a chemical symbol-system, governed by grammatical rules, which scans the environment for antigens. The neural system goes one step further. With our neural adaptation/defence system (NAD) we create an internal image of the world in which we live, as we have perceived it through our senses. Spoken and written language have added to this memory bank: a large part of the perception and interpretation of our environment has come to us by means of oral and written verbal information. Since the 1930's the movement of General Semantics has been studying the many ways in which we perceive, interpret, misconceive and misinterpret the world in which we live.
Speech and language are a phylogenetically young, verbal system for adaptation and defence (VAD). Analogies with man's other systems for adaptation and defence make it very probable that the organisation of language within the neuronal system is a concentrical one. As in the case of the immune and the neural systems the VAD has a firm, genetically based core, which by differentiation has generated pliable surface structures (Chapter 7).
[to continue: click on link in left frame]